![]()
![]()
![]()
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
URL: http://jams.arakmu.ac.ir/article-1-6261-fa.html



2- گروه فیزیولوژی ورزشی، دانشگاه آزاد اسلامی واحد تهران مرکزی، تهران، ایران. ،
3- گروه فیزیولوژی ورزشی، دانشگاه آزاد اسلامی واحد علوم و تحقیقات، تهران، ایران.
مقدمه
عضله اسکلتی نقش مهمی در حفظ وضعیت بدن، حرکت، صحبت کردن، تنفس، تأمین حرکت و نیازهای سوختوسازی دارد. از منظر فیزیولوژیکی، عضله اسکلتی بافتی پویاست که قادر به سازگاری با تحریکات فیزیولوژیکی گوناگون از جمله تمرینات ورزشی است [1]. بافت تاندون امتداد ماتریکس خارج سلولی عضلانی است که به صورت مکانیکی و ساختاری به صورت هماهنگ با عضله با بار مکانیکی سازگاری مییابد [2]. قبلاً تصور میشد تاندون یک بافت غیرفعال از نظر متابولیک است، اما مطالعات اخیر نشان دادهاند تاندون نیز مانند عضله اسکلتی، یک بافت پویاست و فعالیت سوختوسازی آن با بار مکانیکی افزایش مییابد. به دنبال یک ورزش منفرد، تاندون، جذب گلوکز و سنتز کلاژن خود را افزایش میدهد و این حالت دو تا سه روز پس از آن همچنان ادامه مییابد [3].
در حالی که تغییرات مکانیکی و مورفولوژیکی که در پاسخ به تمرینات ورزشی در تاندونها رخ میدهد بهخوبی مستند شده است، درباره سازوکارهای اساسی سلولی و مولکولی که این پاسخها را تنظیم میکنند، اطلاعات زیادی در دسترس نیست. به نظر میرسد که مسیر پیامرسانی خانواده عامل رشدی تغییر شکل دهنده بتا 1 (TGF-β1) نقش اصلی را در سازگاری تاندون با ورزش ایفا میکند. میوستاتین عضو خانواده بزرگ TGF-β است که بیان آن به طور منفی رشد عضله اسکلتی را تنظیم میکند [4]. هم TGF-β1 و هم میوستاتین تکثیر فیبروبلاست تاندون و سنتز کلاژن نوع I را تحریک میکنند [5، 6]. در حالی که به نظر میرسد پیامرسانی TGF-β1 و میوستاتین برای فراخوانی و حفظ فیبروبلاستهای تاندونی در طول تکامل از اهمیت زیادی برخوردار است [5، 7] و مطالعات متعددی وجود دارند که توانایی این سایتوکاینها در القای تکثیر فیبروبلاست و سنتز کلاژن نوع I را در شرایط آزمایشگاهی نشان میدهند [2]، با این حال، هنوز نقش این مسیرهای پیامرسانی در رشد و ترمیم تاندون آزمودنی بالغ، با تردیدهایی روبهروست. در بیشتر مطالعات صورتگرفته در زمینه سازگاریهای ناشی از تمرینات ورزشی، بیشتر خود عضلات اسکلتی مد نظر بوده [8-10] و سازگاریهای تاندونی چندان مورد توجه قرار نگرفته است. بر اساس پیشینه تحقیقاتی موجود، در مطالعات آزمایشگاهی روی فیبروبلاستهای تاندون بالغ و در مطالعات سنتز کلاژن در آزمودنیهای بالغ در پاسخ به تمرین ورزشی، تصور بر این است که TGF-β1 و میوستاتین نقش مهمی در رشد و سازگاری تاندونهای بالغ با تمرین ورزشی ایفا میکنند [11].
ماتریکس متالوپروتئینازها آندوپپتیدازهای وابسته به روی هستند که کلاژن و سایر مولکولهای ساختاری را میشکنند. MMPها برای هموستاز پایه ECM ضروری هستند. MMPها پروتئینازهای اصلی تجزیهکننده کلاژن در ECM به شمار میروند [2]. تاندون یکی از مهمترین اجزایی است که بیان MMP9 در آن رخ میدهد و نشان داده شده است که تمرین ورزشی موجب تحریک سلولهای تاندون برای بیان MMP9 میشود [12]. بیان ژن MMPها توسط سایتوکاینها و مولکولهای پیامرسان مختلف تنظیم میشود که از آن جمله میتوان به TGF-β1 ،IL-1 ،TNF-α و Wnt اشاره کرد، اما اجزای دقیق مسیرهای انتقال سیگنال که بیان MMP به دنبال تمرینات ورزشی را تنظیم میکنند، هنوز به طور کامل آشکار نشده است [2]؛ بنابراین، این مطالعه با هدف بررسی اثر شش هفته تمرین هوازی بر بیان ژنهای TGF-β1، میوستاتین و MMP9 در تاندون عضلات تند و کندانقباض موشهای صحرایی نر نژاد ویستار انجام شد.
مواد و روشها
پژوهش حاضر از نوع تجربی است که به شیوه آزمایشگاهی انجام شد. آزمودنیهای تحقیق حاضر دوازده سر رت نر بالغ نژاد ویستار هشتهفتهای با میانگین وزنی 20±195 گرم بودند که از انستیتو پاستور ایران تهیه شد. رتها در دمای محیطی 3±22 درجه سانتیگراد، رطوبت حدود 45 درصد و چرخه روشنایی /تاریکی 12:12 ساعت نگهداری شدند؛ به طوری که در دسترسی به آب و غذای استاندارد، محدودیتی نداشته باشند. بعد از یک هفته آشناسازی با محیط نگهداری، رتها به روش تصادفی ساده به دو گروه تقسیم شدند: گروه ورزش (تعداد = شش) و گروه کنترل (تعداد = شش)؛ گروه ورزش به مدت شش هفته تمرین هوازی را اجرا کردند.
قبل از اجرای پروتکل تمرینی، تمامی حیوانات به مدت یک هفته با نحوه انجام فعالیت روی نوارگردان آشنا شدند. برنامه آشنایی شامل پنج جلسه راه رفتن و دویدن با سرعت 5 تا 8 متر در دقیقه و شیب صفر درصد به مدت 10 دقیقه بود. برنامه فعالیت بدنی هوازی با افزایش تدریجی سرعت و زمان آغاز شد؛ به طوری که در هفته اول با سرعت 10 متر در دقیقه و به مدت 10 دقیقه، در هفته دوم با سرعت 10 متر در دقیقه و به مدت 20 دقیقه، در هفته سوم با سرعت 15-14 متر در دقیقه و به مدت 20 دقیقه، در هفته چهارم با سرعت 15-14 متر در دقیقه و به مدت 30 دقیقه، در هفته پنجم با سرعت 18-17 متر در دقیقه و به مدت 30 دقیقه و در هفته ششم با سرعت 18-17 متر در دقیقه و به مدت 40 دقیقه بود.
در ابتدا و انتهای هر جلسه تمرینی 3 دقیقه گرم کردن و سرد کردن با سرعت 5-4 متر در دقیقه انجام پذیرفت. به منظور تحریک موشها برای دویدن، از محرک صوتی (ضربه به دیواره نوارگردان) استفاده شد؛ بدینصورت که در جلسات اول، از محرک الکتریکی با ولتاژ کم و همراه با محرک صوتی استفاده شد و پس از شرطی کردن حیوانات به همراه بودن دو محرک، در سایر جلسات به منظور رعایت نکات اخلاقی کار با حیوان آزمایشگاهی، فقط از محرک صوتی استفاده شد [13].
48 ساعت بعد از آخرین جلسه تمرینی، تمامی رتها از طریق تزریق داخل صفاقی مخلوط کتامین (90 میلیگرم / کیلوگرم) و زایلازین (10 میلیگرم / کیلوگرم) بیهوش شدند. سپس، برش در ناحیه شکم و قفسه سینه ایجاد شد و با کشیدن خون به وسیله سرنگ مستقیماً از قلب، حیوانات کشته شدند. سپس، تاندون عضلات نعلی و بازکننده بلند انگشتان پای راست آنها بلافاصله با دقت استخراج و در نیتروژن مایع قرار داده شد و به منظور تجزیه و تحلیل بعدی در دمای منفی 80 درجه سانتیگراد نگهداری شد.
به منظور استخراج RNA تام از بافت تاندون هموژنشده، 1 میلیلیتر ترایزول (Invitrogen, CN 15596018, USA) به 100 میلیگرم بافت اضافه و پس از مخلوط کردن کامل به مدت 15 دقیقه در دمای اتاق انکوبه شد. سپس، 100 میکرولیتر کلروفرم سرد (Merek, CAS 67-66-3 102445, Germany) به نمونه اضافه و به مدت 15 ثانیه مخلوط شدند. محلول در دمای 4 درجه سانتیگراد و با سرعت دوازده هزار دور در دقیقه به مدت 15 دقیقه سانتریفیوژ شد. مایع حاوی RNA به میکروتیوب دیگری انتقال داده شد، سپس 500 میکرولیتر ایزوپروپانول به محلول RNA اضافه شد و 24 ساعت در دمای منفی 20 درجه سانتیگراد نگهداری شد. پس از 24 ساعت، این محلول نیز به مدت 15 دقیقه (دمای 4 درجه سانتیگراد، دوازده هزار دور در دقیقه) سانتریفیوژ شد. مایع رویی با دقت خارج شد و 1 میلیلیتر اتانول 70 درصد خالص سرد به رسوب RNA اضافه شد و سپس به مدت 5 دقیقه (در دمای 4 درجه با 7500 دور در دقیقه) سانتریفیوژ شد.
در ادامه مایع رویی بهدقت خارج شد و رسوب RNA با 100 میکرولیتر ایلوشن بافر (Sigma Aldrich, H5413, Germany) رقیق شد. تمام مراحل استخراج، زیر هود و با مواد و وسایل کاملاً استریل انجام گرفت. در پایان، غلظت و نسبت جذبی نمونهها با استفاده از دستگاه اسپکتوفتومتر (Eppendorf, Germany) ارزیابی شد و نسبت جذبی 260 به 280 بین 1/6-1/8 به عنوان تخلیص مطلوب تعریف شد.
پس از استخراج RNA با خلوص و غلظت بالا، مراحل سنتز cDNA مطابق پروتکل شرکت سازنده (high-capacity cDNA reverse transcription kit) انجام شد. ابتدا RNA، پرایمر و آب با هم ترکیب شدند و محلول به مدت 5 دقیقه در دمای 65 درجه سانتیگراد انکوبه شد. سپس محلول به مدت، 2 دقیقه روی یخ قرار گرفت. پس از آن enzyme mix و reaction buffer به محلول اضافه شدند. محلول در سه مرحله متوالی انکوبه شد: مرحله اول، به مدت 10 دقیقه در دمای 25 درجه سانتیگراد؛ مرحله دوم، به مدت 30 دقیقه در دمای 42 درجه سانتیگراد و مرحله سوم، به مدت 5 ثانیه در دمای 8 درجه سانتیگراد انجام شد و درنهایت cDNA سنتز شد و در دمای منفی 80 درجه سانتیگراد ذخیره شد. تمام مراحل انجام کار روی یخ، زیر هود و با استفاده از وسایل RNase free انجام شد.
برای اندازهگیری سطوح بیان mRNA از روش کمی Real time-PCR استفاده شد. در ابتدای کار میزان غلظت بهینه cDNA و همچنین پرایمرهای مربوط به هر ژن با استفاده از آزمایش سریال غلظت برای هرکدام به طور جداگانه مشخص شد؛ به طوری که کمترین میزان دایمر و بهترین Ct مشاهده شود. Real time-PCR با استفاده از RealQ Plus 2x Master Mix Green شرکت AMPLIQON و با استفاده از غلظت 250 نانوگرم از cDNA انجام گرفت.
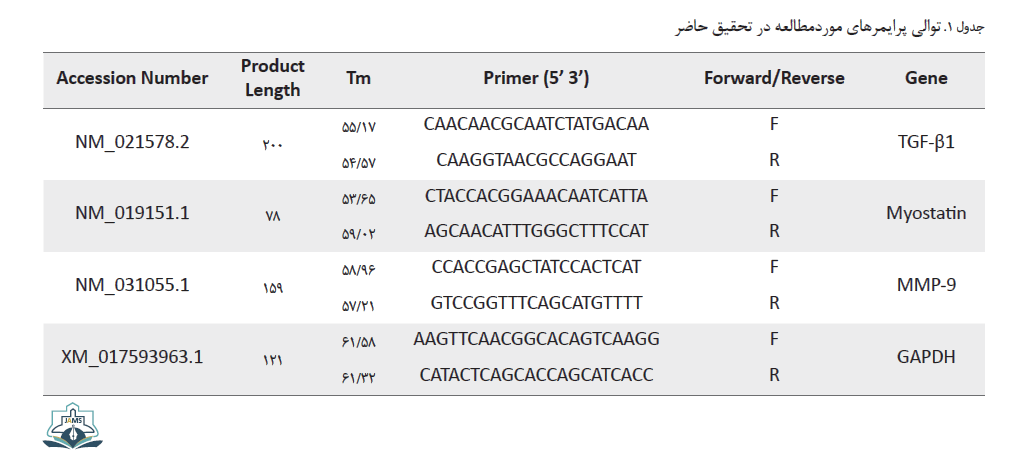
توالی پرایمرهای مربوط به متغیرهای موردمطالعه بر اساس اطلاعات این ژنها در بانک ژنی NCBI توسط شرکت پیشگام (ایران) طراحی شد (جدول شماره 1).
برنامه Real time-PCR شامل واسرشت اولیه در دمای 95 درجه سانتیگراد و به مدت10 دقیقه، واسرشت در هر سیکل PCR در دمای 95 درجه سانتیگراد به مدت 15 ثانیه و با توجه به دمای اتصال پرایمرها هر سیکل به مدت 30 ثانیه (چهل سیکل) در نظر گرفته شد. نمودار دمای ذوب جهت بررسی صحت واکنشهای انجامشده و به صورت اختصاصی برای هر ژن و در هر بار از واکنش به همراه نمودار کنترل منفی جهت بررسی وجود آلودگی در هر واکنش مورد ارزیابی قرار گرفت. از ژن گلیسرآلدهید 3 فسفات دهیدروژناز به عنوان ژن کنترل استفاده شد و میزان بیان ژن موردنظر با فرمول .JPG) محاسبه شد [14].
محاسبه شد [14].
برای بررسی طبیعی بودن توزیع دادهها از آزمون کولموگروف اسمیرنف استفاده شد. به منظور تحلیل دادهها از آزمون تی مستقل استفاده شد و معنیداری بین متغیرها در سطح p≤0/05 مورد توجه قرار گرفت. برای تجزیه و تحلیل دادهها، از نرمافزار SPSS نسخه 16 و برای ترسیم نمودارها از نرمافزار Excel 2016 استفاده شد.
یافتهها
نتایج نشان داد بین مقادیر ژن TGF-β1 عضلات EDL و نعلی گروه تمرین هوازی در مقایسه با گروه کنترل تفاوت معنیداری وجود دارد. بیان ژن TGF-β1 عضلات EDL و نعلی در گروه تمرین هوازی در مقایسه با گروه کنترل به طور معنیداری افزایش یافت (عضله EDL: 0/09 ± 0/63 در مقایسه با 0/04 ± 0/14، 0/001 ≥P و عضله سولئوس: 0/08 ± 0/36 در مقایسه با 0/04 ± 0/17، 0/001≥P) (تصویر شماره 1).
.png)
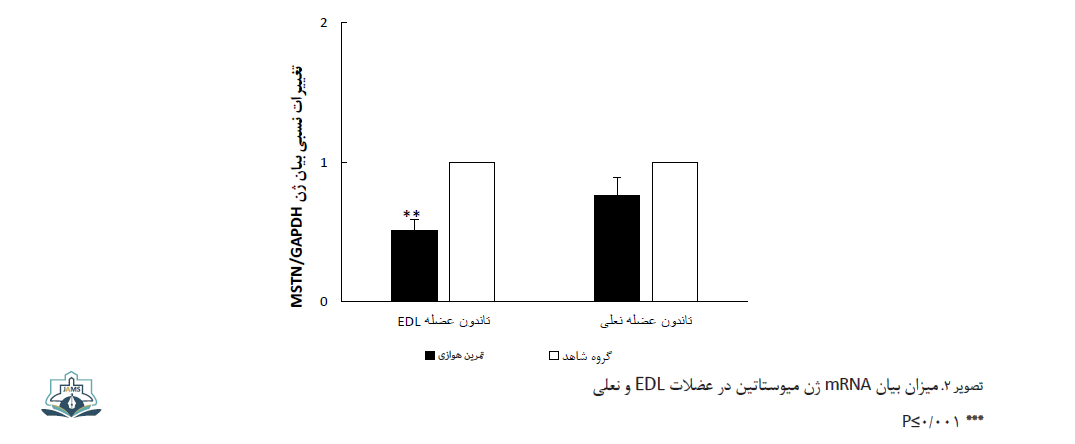
نتایج آزمون تی مستقل نشان داد بیان ژن میوستاتین تنها در عضله EDL گروه تمرین هوازی در مقایسه با گروه کنترل به طور معنیداری کاهش یافته است (عضله EDL: 0/1 ± 0/3 در مقایسه با 0/07 ± 0/58، 0/001≥P و عضله سولئوس: 0/09 ± 0/20 در مقایسه با 0/05 ± 0/29، 0/001≥P) (تصویر شماره 2).
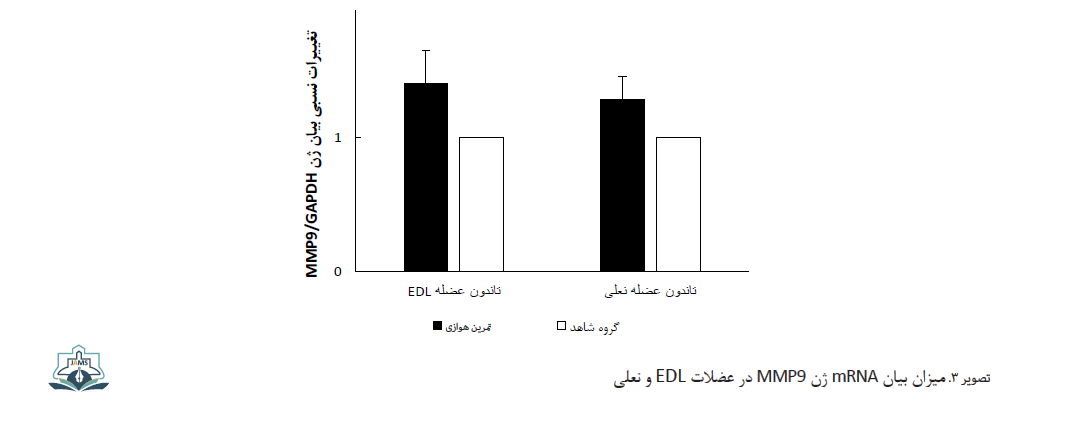
نتایج آزمون تی مستقل نشان داد تفاوت معنیداری بین بیان ژن MMP9 در عضلات EDL و نعلی بین دو گروه تمرین هوازی و گروه کنترل وجود ندارد (عضله EDL: 0/08 ± 0/21 در مقایسه با 0/02 ± 0/15، 0/05<P و عضله سولئوس: 0/07 ± 0/27 در مقایسه با 0/1 ± 0/19، 0/05<P) (تصویر شماره 3).
بحث
مطالعه حاضر با هدف بررسی اثر شش هفته تمرین هوازی بر بیان ژنهای TGF-β1، میوستاتین و MMP9 در تاندون عضلات تند و کندانقباض موشهای نر ویستار انجام شد. نتایج مطالعه حاضر نشان داد به دنبال شش هفته برنامه تمرین هوازی دویدن روی تردمیل، میزان بیان mRNA ژن TGF-β1 در هر دو عضله EDL و نعلی به شکل معنیداری افزایش مییابد. همچنین، شش هفته برنامه تمرین هوازی موجب کاهش معنیدار میزان بیان mRNA ژن میوستاتین در عضله EDL شد. میزان بیان ژن میوستاتین در عضله نعلی نیز کاهش یافت، ولی از لحاظ آماری معنیدار نبود. به علاوه، بیان mRNA ژن MMP9 در عضلات EDL و نعلی افزایش غیرمعنیدار نشان داد.
مرور متون پژوهشی حاکی از آن است که بیشتر مطالعات صورتگرفته در این حوزه به پاسخ و سازگاریهای عضلات اسکلتی با تمرین ورزشی پرداختهاند و بافت تاندون چندان مورد توجه قرار نگرفته است. این امر مقایسه یافتههای مطالعه حاضر با پژوهشهای پیشین را دشوار میکند. یکی از یافتههای مطالعه حاضر، افزایش معنیدار بیان mRNA ژن TGF-β1 در هر دو عضله EDL و نعلی است. هنیمیر و همکاران افزایش سطوح mRNA ژنهای TGF-β1 و کلاژنهای نوع I و III در عضله دوقلوی میانی و تاندون آشیل به دنبال انجام انقباضات ایزومتریک، کانسنتریک و اکسنتریک از طریق تحریک عصب سیاتیک به مدت چهار روز را گزارش کردهاند [15]. همچنین، در تحقیق دیگری نشان داده شد که انجام تمرین استقامتی طولانیمدت باعث افزایش قابل توجهی در mRNA ژن TGF-β1 در عضله نعلی شده است [8].
سازوکارهایی که در طول تمرینات ورزشی بار مکانیکی را به افزایش سطوح کلاژن در واحد عضله تاندون مرتبط میکنند، هنوز به طور کامل شناخته نشدهاند. نشان داده شده است که وضعیت آنتیاکسیدانی عضله بر بیان TGF-β1 اثرگذار است. سزارکوسکا پازک و همکاران گزارش کردند که افزایش استرس اکسیداتیو از طریق افزایش بیان NADPH اکسیداز و کاهش فعالیت آنزیم سوپراکسید دیسموتاز، بیان TGF-β1 را افزایش میدهد [16]. درحقیقت، TGF-β1 سایتوکاینی است که خاصیت آنتیاکسیدانی دارد و در پاسخ به التهاب عضله افزایش مییابد.
انجام تمرینات ورزشی در درازمدت قدرت آنتیاکسیدانی عضله را افزایش میدهد [8]. شواهد موجود نشاندهنده آن است که TGF-β1 واسطه اصلی القای سنتز کلاژن در فیبروبلاستها از طریق بار مکانیکی بوده [15] و چنین نقشی برای TGF-β1 در تاندونها نیز پیشنهاد شده است [17]. افزایش بالای 453 درصدی میزان بیان mRNA ژن TGF-β1 در تاندون عضله EDL در برابر افزایش 203درصدی آن در تاندون عضله نعلی به دنبال تمرینات هوازی که در مطالعه حاضر مشاهده شد، به احتمال زیاد حاکی از درگیر شدن عضلات تندانقباض در تمرینات هوازی است که در اعمال معمول روزانه کمتر مورد استفاده قرار میگیرد.
میوستاتین علاوه بر اینکه اندازه، نوع و انقباضپذیری عضله را کنترل میکند، ترکیب ECM عضله و تاندون را احتمالاً از طریق القای بیان کلاژن نوع I تنظیم میکند (مندیاس و همکاران 2008). با این حال، مندیاس و همکاران نشان دادند که غیرفعالسازی ژنتیکی میوستاتین در رتها ضمن افزایش حجم عضلانی، تأثیر منفی بر ویژگیهای مکانیکی تاندون ندارد [18]. در مطالعه حاضر، تمرینات هوازی باعث کاهش سطوح پایه بیان mRNA ژن میوستاتین در تاندون عضله EDL به شکل معنیدار و تاندون عضله نعلی به شکل غیرمعنیدار شد. کاهش سطوح سرمی و عضلانی میوستاتین به دنبال تمرینات ورزشی در بیشتر مطالعات گزارش شده است [9، 10، 19]. با این حال، بر اساس جستوجوهای ما، تنها مطالعهای که تغییرات میوستاتین در اثر سازگاری ورزشی در تاندون را اندازهگیری کرده است، عدم تغییر سطوح بیان mRNA ژن میوستاتین در تاندون آشیل و کاهش آن در عضله دوقلوی میانی به دنبال انجام تمرینات مقاومتی کوتاهمدت را نشان داده است [15]. از دلایل تفاوت در یافتهها میتوان به مدت و نوع پروتکل تمرین ورزشی اشاره کرد. در مطالعه مذکور از تحریک عصب سیاتیک به منظور اعمال برنامه تمرین مقاومتی در رتها استفاده شده بود که تنها چهار روز ادامه داشت، در حالی که برنامه تمرین هوازی استفادهشده در مطالعه حاضر دویدن روی تردمیل به مدت شش هفته بود.
به طور جالب، گزارش شده است که نوع ورزش عامل اصلی تعیینکننده برای فراوانی رونویسی RNA میوستاتین به شمار نمیرود [20]. عطارزاده حسینی و همکاران کاهش سطوح مایوستاتین سرمی در زنان را به دنبال هشت هفته تمرینات مقاومتی با شدت بالا گزارش کردند [19]. هیتل و همکاران تأثیر شش ماه تمرین هوازی متوسط را بر مقادیر میوستاتین عضلانی و پلاسمایی مردان میانسال مورد بررسی قرار داده و کاهش متوسط 37درصدی میوستاتین عضلانی و پلاسمایی را بعد از تمرین گزارش کردند [9].
در تحقیقی دیگر، ماتساکاس و همکاران نتایج متفاوتی را در بیان ژن میوستاتین در پاسخ به تمرین استقامتی در بافتهای مختلف گزارش کردند. در عضله دوقلوی موشهای صحرایی تمرینکرده در بیان ژن میوستاتین کاهش 65درصدی گزارش شد، در حالی که در عضله پهن خارجی این کاهش متوسط و به مقدار 49 درصد بود و در عضله نعلی بین موشهای تمرینکرده و بیتمرین تغییری مشاهده نشد [10]. درحقیقت، شواهد زیادی از این ایده حمایت میکنند که تنظیم میوستاتین با توجه به نوع تارهای عضلانیای که وابستگی دارد، بهشدت با ایزوفرم IIb زنجیره سنگین میوزین عضله ارتباط دارد [21] و غلظت بالای پروتئین میوستاتین در عضله تندانقباض در مقایسه با تارهای کندانقباض مشاهده شده است [22]. این گزارشات میتواند نتایج حاصل از مطالعه حاضر مبنی بر کاهش بالای 49درصدی بیان mRNA ژن میوستاتین در عضله EDL در مقایسه با کاهش اندک 24 درصدی بیان آن در عضله نعلی به دنبال تمرینات هوازی را توجیه کنند.
MMP9 همراه با دیگر عضو زیرگروه MMP ژلاتیناز، یعنی MMP2، کلاژنهای آسیبدیده و تغییر ماهیتداده نوع IV، VII و X را تجزیه میکند [23، 24]. بنابراین، به نظر میرسد که تمرینات ورزشی باعث آسیب به بافت کلاژنی عضلات اسکلتی و تاندونها شده و افزایش سطوح MMP9 در جهت حذف این عناصر آسیبدیده از بافت رخ میدهد. علاوه بر این، گزارش شده است که وجود MMPها برای القای آنژیوژنز ناشی از تمرینات ورزشی در عضلات اسکلتی ضروری است [25]. در مطالعه حاضر، تمرین ورزشی باعث افزایش بیان MMP9 در عضلات EDL (40 درصد افزایش) و نعلی (28 درصد افزایش) شد، اما این تغییرات از لحاظ آماری معنیدار نبود. همسو با یافتههای مطالعه حاضر، رولمن و همکاران افزایش غیرمعنیدار میزان بیان MMP9 در عضله پهن خارجی انسان را به دنبال تمرین هوازی گزارش کردهاند [26].
کاهش سطوح میوستانین به دنبال تمرین هوازی که در مطالعه حاضر مشاهده شد، میتواند به عنوان سازوکاری برای افزایش نسبی بیان mRNA ژن MMP9 در بافت تاندون در نظر گرفته شود. مندیاس و همکاران افزایش 141درصدی بیان ژن MMP9 را در بافت تاندون رتهای فاقد میوستاتین (رتهایی که میوستاتین آنها از طریق ژنتیکی غیرفعال شده بود) گزارش کردند [18]. به علاوه، نشان داده شده است که فعالسازی آبشار پیامرسانی ERK1/2 و JNK-NF-κB از طریق ROS برای تنظیم مثبت / فعالسازی MMP9 و مهاجرت سلولی ایجادشده توسط TGF-β1 ضروری است [27]. بر اساس این گزارش، افزایش سطوح TGF-β1 میتواند سازوکار دیگری برای توجیه افزایش نسبی بیان mRNA ژن MMP9 در بافت تاندون در این مطالعه باشد.
نتیجهگیری
تمرین هوازی باعث تنظیم مثبت سطوح پایه ژن TGF-β1 و تنظیم منفی سطوح پایه ژن میوستاتین در تاندون عضلات تند و کند انقباض میشود؛ و این اثرات در تاندون عضله تندانقباض به طور قابل توجهی بیشتر است. به نظر میرسد که تمرین هوازی میتواند بیان ژنهای درگیر در تنظیم کلاژن بافت تاندون را در یک روش وابسته به نوع عضله تعدیل کند.
ملاحظات اخلاقی
پیروی از اصول اخلاق پژوهش
همه مراحل مربوط به کار با حیوانات با توجه به دستورالعمل اخلاقی و مجوز معاونت پژوهشی دانشگاه آزاد اسلامی واحد تهران مرکزی با شماره IR.IAU.PS.REC.1398.296 انجام شده است.
حامی مالی
این مقاله از رساله دکتری نویسنده دوم، در گروه فیزیولوژی ورزشی، دانشکده تربیتبدنی و علوم ورزشی دانشگاه آزاد اسلامی واحد تهران مرکزی استخراج شده است.
مشارکت نویسندگان
تعریف موضوع و بیان مسئله، نگارش متن و بازبینی: تمام نویسندگان؛ روش پژوهش و تحلیل دادهها: قاسم محمدنژاد.
تعارض منافع
نویسندگان مقاله هیچگونه تعارضی در منافع اعلام نکردند.
تشکر و قدردانی
نویسندگان مقاله از جناب دکتر رضا قراخانلو، گروه تربیتبدنی و علوم ورزشی، دانشکده علوم انسانی و دانشگاه تربیت مدرس به خاطر همفکری در طراحی موضوع و بیان مسئله تقدیر و تشکر میکنند.
Reference
1.Schiaffino S, Reggiani C. Molecular diversity of myofibrillar proteins: Gene regulation and functional significance. Physiol Rev. 1996; 76(2):371-423. [DOI:10.1152/physrev.1996.76.2.371] [PMID]
2.Davis ME, Gumucio JP, Sugg KB, Bedi A, Mendias CL. MMP inhibition as a potential method to augment the healing of skeletal muscle and tendon extracellular matrix. J Appl Physiol. 2013; 115(6):884-91. [DOI:10.1152/japplphysiol.00137.2013] [PMID] [PMCID]
3.Kjær M, Langberg H, Heinemeier K, Bayer M, Hansen M, Holm L, et al. From mechanical loading to collagen synthesis, structural changes and function in human tendon. Scand J Med Sci sports. 2009;19(4):500-10. [DOI:10.1111/j.1600-0838.2009.00986.x] [PMID]
4.McPherron AC, Lawler AM, Lee S-J. Regulation of skeletal muscle mass in mice by a new TGF-p superfamily member. Nature. 1997; 387(6628):83-90. [DOI:10.1038/387083a0] [PMID]
5.Mendias CL, Bakhurin KI, Faulkner JA. Tendons of myostatin-deficient mice are small, brittle, and hypocellular. Proc Natl Acad Sci. 2008; 105(1):388-93. [DOI:10.1073/pnas.0707069105] [PMID] [PMCID]
6.Mendias CL, Gumucio JP, Bakhurin KI, Lynch EB, Brooks SV. Physiological loading of tendons induces scleraxis expression in epitenon fibroblasts. J Orthop Res. 2012; 30(4):606-12. [DOI:10.1002/jor.21550] [PMID] [PMCID]
7.Pryce BA, Watson SS, Murchison ND, Staverosky JA, Dünker N, Schweitzer R. Recruitment and maintenance of tendon progenitors by TGFβ signaling are essential for tendon formation. Dev. 2009; 136(8):1351-61. [DOI:10.1242/dev.027342] [PMID] [PMCID]
8.Czarkowska-Paczek B, Zendzian-Piotrowska M, Bartlomiejczyk I, Przybylski J, Gorski J. The effect of acute and prolonged endurance exercise on transforming growth factor-beta1 generation in rat skeletal and heart muscle. J physiol pharmacol. 2009; 60(4):157-62. [PMID]
9.Hittel DS, Axelson M, Sarna N, Shearer J, Huffman KM, Kraus WE. Myostatin decreases with aerobic exercise and associates with insulin resistance. Med Sci Sports Exerc. 2010; 42(11):2023. [DOI:10.1249/MSS.0b013e3181e0b9a8] [PMID] [PMCID]
10.Matsakas A, Friedel A, Hertrampf T, Diel P. Short‐term endurance training results in a muscle‐specific decrease of myostatin mRNA content in the rat. Acta Physiol Scand. 2005; 183(3):299-307. [DOI:10.1111/j.1365-201X.2005.01406.x] [PMID]
11.Gumucio JP, Sugg KB, Mendias CL. TGF-β superfamily signaling in muscle and tendon adaptation to resistance exercise. Exerc sport Sci Rev. 2015; 43(2):93. [DOI:10.1249/JES.0000000000000041] [PMID] [PMCID]
12.Koskinen SO, Heinemeier KM, Olesen JL, Langberg H, Kjaer M. Physical exercise can influence local levels of matrix metalloproteinases and their inhibitors in tendon-related connective tissue. J Appl Physiol. 2004; 96(3):861-4. [DOI:10.1152/japplphysiol.00489.2003] [PMID]
13.Sadighi A AA, Azarbayjani MA, Barari AR. [Effect of aerobic exercise on some factors of cardiac apoptosis in male rats (Persian)]. Feyz. 2019; 23(5):495-502. http://feyz.kaums.ac.ir/article-1-3792-fa.html
14.Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001; 29(9):e45-e. [DOI:10.1093/nar/29.9.e45] [PMID] [PMCID]
15.Heinemeier K, Olesen J, Haddad F, Langberg H, Kjær M, Baldwin K, et al. Expression of collagen and related growth factors in rat tendon and skeletal muscle in response to specific contraction types. J Physiol. 2007; 582(3):1303-16. [DOI:10.1113/jphysiol.2007.127639] [PMID] [PMCID]
16.Czarkowska-Paczek B, Bartlomiejczyk I, Przybylski J. The serum levels of growth factors: PDGF, TGF-beta and VEGF are increased after strenuous physical exercise. J Physiol Pharmacol. 2006; 57:189-97. [PMID]
17.Yang G, Crawford RC, Wang JH. Proliferation and collagen production of human patellar tendon fibroblasts in response to cyclic uniaxial stretching in serum-free conditions. J Biomech. 2004; 37(10):1543-50. [DOI:10.1016/j.jbiomech.2004.01.005] [PMID]
18.Mendias CL, Lynch EB, Gumucio JP, Flood MD, Rittman DS, Van Pelt DW, et al. Changes in skeletal muscle and tendon structure and function following genetic inactivation of myostatin in rats. J Physiol. 2015; 593(8):2037-52. [DOI:10.1113/jphysiol.2014.287144] [PMID] [PMCID]
19.Attarzadeh Hosseini SR, Moeinnia N, Motahari Rad M. The effect of two intensities resistance training on muscle growth regulatory myokines in sedentary young women. Obes Med. 2017; 5:25-8. [DOI:10.1016/j.obmed.2017.01.004]
20.Matsakas A, Bozzo C, Cacciani N, Caliaro F, Reggiani C, Mascarello F, et al. Effect of swimming on myostatin expression in white and red gastrocnemius muscle and in cardiac muscle of rats. Exp Physiol. 2006; 91(6):983-94. [DOI:10.1113/expphysiol.2006.033571] [PMID]
21.Carlson CJ, Booth FW, Gordon SE. Skeletal muscle myostatin mRNA expression is fiber-type specific and increases during hindlimb unloading. Am J Physiol-Regul Integr Comp Physiol. 1999; 277(2):R601-R6. [DOI:10.1152/ajpregu.1999.277.2.R601] [PMID]
22.Wehling M, Cai B, Tidball JG. Modulation of myostatin expression during modified muscle use. FASEB J. 2000; 14(1):103-10. [DOI:10.1096/fasebj.14.1.103] [PMID]
23.Kim J, Lee J. Matrix metalloproteinase and tissue inhibitor of metalloproteinase responses to muscle damage after eccentric exercise. J Exerc Rehabil. 2016; 12(4):260-5. [DOI:10.12965/jer.1632640.320] [PMID] [PMCID]
24.Monaco S, Sparano V, Gioia M, Sbardella D, Di Pierro D, Marini S, et al. Enzymatic processing of collagen IV by MMP‐2 (gelatinase A) affects neutrophil migration and it is modulated by extracatalytic domains. Protein Sci. 2006; 15(12):2805-15. [DOI:10.1110/ps.062430706] [PMID] [PMCID]
25.Haas T, Milkiewicz M, Davis S, Zhou A, Egginton S, Brown M, et al. Matrix metalloproteinase activity is required for activity-induced angiogenesis in rat skeletal muscle. Am J Physiol-Regul Integr Comp Physiol. 2000; 279(4):H1540-H7. [DOI:10.1152/ajpheart.2000.279.4.H1540] [PMID]
26.Rullman E, Norrbom J, Strömberg A, Wågsäter D, Rundqvist H, Haas T, et al. Endurance exercise activates matrix metalloproteinases in human skeletal muscle. J Appl Physiol. 2009; 106(3):804-12. [DOI:10.1152/japplphysiol.90872.2008] [PMID]
27.Hsieh HL, Wang HH, Wu WB, Chu PJ, Yang CM. Transforming growth factor-β1 induces matrix metalloproteinase-9 and cell migration in astrocytes: roles of ROS-dependent ERK-and JNK-NF-κB pathways. J neuroinflammation. 2010; 7(1):88. [DOI:10.1186/1742-2094-7-88] [PMID] [PMCID]
| بازنشر اطلاعات | |
 |
این مقاله تحت شرایط Creative Commons Attribution-NonCommercial 4.0 International License قابل بازنشر است. |